Яды — высокоточное оружие: компьютерное исследование природных нейротоксинов
10 декабря 2013
Яды — высокоточное оружие: компьютерное исследование природных нейротоксинов
- 5565
- 2
- 11
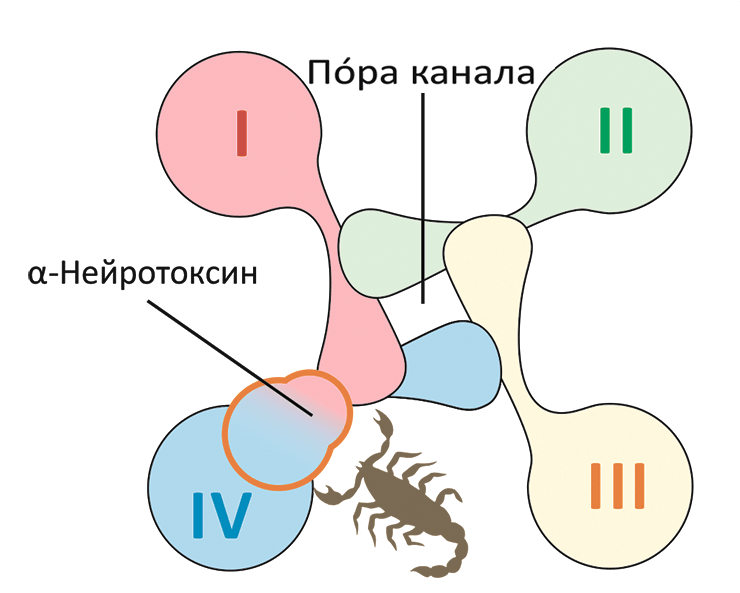
Потенциал-чувствительный натриевый канал имеет форму диафрагмы фотоаппарата, где пóровая часть одного домена канала взаимодействует с потенциал-чувствительной частью другого домена. α-Нейротоксины, по-видимому, отражают это строение в своей модульной архитектуре.
[23], рисунок адаптирован
-
Авторы
-
Редакторы
Биологическая эволюция — общая форма существования живой материи. При детальном рассмотрении оказывается, что виды почти никогда не эволюционируют поодиночке: обычно в этом принимают участие их экологические партнеры, и изменение происходит в парах паразит—хозяин или хищник—жертва. Более того, коэволюцию часто можно проследить на молекулярном уровне, когда один вид совершенствует систему нападения, а второй вслед за ним — систему защиты (и наоборот). Параллель с холодной войной здесь настолько очевидна, что соответствующий феномен даже получил название эволюционной «гонки вооружений». Примером наиболее эффективных и элегантных систем нападения служат животные яды, содержащие в своем составе нейротоксины — вещества, воздействующие на нервную систему и мышцы жертвы.
Несмотря на детальную проработку теории эволюции профессионалами и многочисленные примеры, вошедшие в учебники и популярную литературу [1], воплощение эволюционного процесса зачастую настолько причудливо, что исследование изменений в живых организмах может быть интересно далеко не только ученым-теоретикам. Известно, что у эволюции нет воли и цели: живые организмы меняются, чтобы оставить максимальное потомство, а не чтобы достигнуть абстрактного совершенства [2]. Так, эволюционно прогрессивным признаком считается то, что позволяет лучше приспособиться к окружающим условиям в данный момент, а не красота или сложность устройства сами по себе.
Например, антарктическая ледяная рыба утратила гены гемоглобина из-за того, что условия ее обитания — нулевая или даже отрицательная (!) температура воды, растворимость кислорода в которой достаточно высокая, чтобы обеспечивать ткани этим газом просто за счет диффузии и усиления кровотока. Можно сказать, что в этом случае рыба «экономит» на гемоглобине [3]. Другие рыбы, которые живут в пещерах и никогда не выплывают на белый свет или обитают на очень больших глубинах, почти всегда за ненадобностью теряют зрение. Степень упрощения паразитических организмов вообще поражает воображение, причем в случае бактерий это приводит к кардинальному упрощению всего генетического аппарата: например, геном микоплазмы содержит всего около 500 генов, а у облигатного внутриклеточного симбионта Candidatus Carsonella ruddii и вовсе около 180 генов [4]. И всё перечисленное — прогрессивные признаки, поскольку они позволили упомянутым организмам максимально адаптироваться к обстановке, в которой они обитают. Более известный (и более «благородный») пример — эволюция человека, где развитие пошло по пути усложнения нервной системы и способности к коммуникации, а также к становлению богатой культуры, современное состояние которой мы имеем удовольствие ежедневно наблюдать вокруг. Одновременно возросла роль полового отбора и образования семей [5], — и все это тоже проделки эволюции [6].
В нашей статье речь пойдет об особой разновидности эволюционного процесса, которую можно назвать «гонкой вооружений» и в которой участвуют две стороны: нападающая и обороняющаяся (или «догоняющая» и «убегающая»). Чрезвычайно любопытно наблюдать «гонку» на молекулярном уровне. Поскольку изменение одной стороны согласовано с изменением второй, то и гонку вооружений иногда можно довольно подробно проследить по «молекулярной летописи» процессов, происходящих внутри живых организмов.
Уроки холодной войны
Вторая половина XX века, кроме замечательных открытий в молекулярной биологии, была ознаменована холодной войной — глобальной конфронтацией между социалистическим и капиталистическим строями. Это противостояние породило эффект гонки вооружений — процесс борьбы за военное превосходство с положительной обратной связью, вызвавший гипертрофию военно-промышленных комплексов СССР и США.
Похожий эффект наблюдается и в процессе коэволюции двух видов, связанных друг с другом в системе «хищник—жертва» или «паразит—хозяин»: изменение одного из видов неизбежно влечет изменение второго для сохранения паритета или получения превосходства. Хорошо известный пример эволюционной гонки вооружений — газель и гепард, каждому из которых приходится бегать из поколения в поколение все быстрее и быстрее, чтобы одному ускользнуть из пасти, а другому — остаться сытому.
Эволюционная гонка вооружений, или принцип Черной королевы
Подчеркивая параллель с холодной войной, биологи ввели понятие эволюционной гонки вооружений (англ. evolutionary arms race), приписывая этому процессу роль важнейшей движущей силы эволюции [7]. Более строго это понятие сформулировано в форме «принципа Черной Королевы», утверждающего, что в коэволюционных отношениях каждому из видов приходится «бежать со всех ног, чтобы только остаться на том же месте» (эта цитата из «Алисы в Зазеркалье» удачно передает суть принципа). Прямое экспериментальное подтверждение принципа Черной королевы получено на микроскопических рачках дафниях и их паразитических бактериях: «воскресив» из донного ила несколько поколений тех и других, биологи показали, что самые опасные для дафний паразиты — современные им, тогда как «прошлые» и «будущие» популяции бактерий заражали рачков с меньшей эффективностью [8].
Око за око, ген за ген
Хорошим примером эволюционной гонки вооружений являются растения с их паразитами. Во многих случаях наблюдается строгое соответствие системы устойчивости растения системе вирулентности его паразита. Более того, это соответствие сводится всего лишь к двум генам: гену растения, позволяющему противостоять заражению (гену резистентности, R) и гену паразита, необходимому для инфекции (гену авирулентности, Avr). Такие межвидовые отношения получили название «ген за ген» (англ. gene-for-gene) [9]. Растения, содержащие ген R, оказываются устойчивыми по отношению к паразитам с соответствующим геном Avr. Как правило, гены резистентности растений кодируют белки-рецепторы, регистрирующие появление паразита. Теперь паразит стремится изменить свой Avr-ген так, чтобы ускользнуть от узнавания рецептором растения. И наоборот, растение изменяет свой R-ген, чтобы по-прежнему детектировать заражение.
Вариация фаз
Одно из наиболее прогрессивных эволюционных приобретений высших позвоночных — развитая иммунная система, основанная на принципе комбинаторики и позволяющая противостоять практически любому чужеродному организму [10], [11]. Однако бактерии и вирусы на сдаются, им тоже есть что предъявить из своего арсенала. Хорошо известный пример — постоянно мутирующий вирус гриппа, к которому необходимо получать всё новые и новые вакцины [12]. Другой, не менее поразительный, пример — явление «смены фаз» у бактерий, заключающееся в случайном изменении фенотипа с высокой частотой, намного превышающей частоту обычных мутаций. Так, сальмонеллы используют этот прием для замены белка флагеллина, из которого построены бактериальные жгутики и который служит сигналом для запуска иммунологических реакций. Только у хозяина разовьется иммунный ответ, как сальмонелла меняет тип флагеллина и ускользает [13]!
Ядовитый арсенал
Пожалуй, самым эффективным средством как нападения, так и защиты служат яды, которые в процессе эволюции научились вырабатывать разнообразнейшие животные: многие кишечнополостные, членистоногие, моллюски, хордовые и другие (рис. 1). Присутствующие в ядах молекулы называют токсинами, а в том частном (но распространенном) случае, когда мишенью их действия служит нервная система и/или мышцы, — нейротоксинами.

Рисунок 1. Разнообразие ядовитых животных. На филогенетическом дереве многоклеточных отмечены типы животных, среди представителей которых встречаются ядовитые. Вокруг дерева расположены фотографии ядовитых животных (сверху — позвоночных, слева — членистоногих, слева внизу — иглокожих, справа вверху — моллюсков, справа внизу — губок, кишечнополостных и некоторых червей).
Состав яда животных различен: если, допустим, в яде пчелы присутствуют всего два основных компонента — мембраноактивный пептид мелиттин и гидролизующий липиды фермент фосфолипаза A2, — то в яде пауков, скорпионов, морских анемон и конусов, а также змей содержатся десятки, а иногда сотни или даже тысячи компонентов различной химической природы. Наблюдаемое в ядах разнообразие компонентов одного структурного типа сегодня принято описывать термином «эволюционно отредактированная комбинаторная библиотека» [14]. Эти молекулы обладают различной эффективностью и специфичностью в отношении разных рецепторов, а результирующая смесь токсинов эффективна в отношении широкого круга мишеней.
Эволюционное преимущество богатого арсенала — способность «следовать» за жертвой: если мишень действия основного компонента яда начнет ускользать (например, рецептор мутирует), в яде с большой вероятностью обнаружится слабо представленный, но более активный по отношению к новой форме рецептора токсин, и теперь уже ему суждено будет стать основой «вооружения» у будущих поколений ядовитых хищников.
«Всенепременнейше захватите телеграф!»
Как уже было сказано, частой мишенью действия ядов является нервная система, а поэтому основа арсенала многих ядовитых гадов — нейротоксины. Связано это, по-видимому, с тем, что нарушить работу сложной системы проще всего, уничтожив коммуникацию — что было отмечено еще в работах Ленина, рекомендовавшего в случае захвата власти в первую очередь взять под контроль телефон, телеграф и железнодорожные станции. «Телеграфом» нашего организма является нервная сеть, передача сообщений в которой основана на феномене потенциала действия, распространяющегося вдоль мембран нейронов. В состоянии покоя поддерживается определенная разница концентраций ионов (Na+, K+, Ca2+, Cl−) внутри нейронов и снаружи, и формируется потенциал покоя [15]. Рабочими элементами потенциала действия служат ионные каналы — мембранные белки, пропускающие те или иные ионы по команде. В частности, начинается потенциал действия с активации потенциал-чувствительных натриевых (Na+) каналов (ПЧНК), открывающихся в ответ на уменьшение трансмембранной разности потенциалов (деполяризацию). Интересно, что появление ПЧНК в эволюции животных тесно связано с дифференцировкой тканей и обособлением нервной системы. О них и поговорим подробнее, оставив другие каналы за рамками нашей статьи.
Каждый ПЧНК состоит из очень длинной полипептидной цепи (около двух тысяч остатков аминокислот), которая представлена четырьмя похожими повторами (рис. 2). При этом в пространстве канал составлен из пяти частей (или доменов). В образовании единственного порового домена (ПД) участвуют все повторы полипептидной цепи; в его центре формируется селективная для ионов Na+ пора. Четыре — по одному от каждого повтора — потенциал-чувствительных домена (ПЧД) расположены вокруг центрального ПД [16]. Функция ПЧД — реагировать на изменение мембранного потенциала и передавать команду на открытие поры.

Рисунок 2. Строение потенциал-чувствительных натриевых каналов (ПЧНК). а — ПЧНК состоят из четырех похожих повторов (I–IV). Каждый повтор содержит шесть трансмембранных α-спиралей (S1—S6). Четыре спирали (S1—S4) формируют потенциал-чувствительный домен (ПЧД). Единственный поровый домен (ПД) образован восемью спиралями, по две (S5—S6) от каждого повтора. Четвертая спираль в каждом ПЧД (S4) положительно заряжена и выполняет роль сенсора потенциала. С внеклеточной стороны с ПЧД-IV и ПЧД-II связываются α- и β-токсины скорпионов. б — В пространстве повторы ПЧНК располагаются вокруг общей оси. в — Центральный ПД канала окружен ПЧД. Участок между спиралями S5 и S6 содержит «селективный фильтр», опосредующий избирательное пропускание ионов Na+.
ПЧНК являются ключевым компонентом передачи нервных импульсов, а значит и работы нервной системы вообще. Очевидно, именно этим обусловлено существование огромного числа токсинов, связывающихся с ПЧНК и так или иначе нарушающих их работу (рис. 3). В свою очередь, нарушение работы ПЧНК приводит к параличу, судорогам, а смерть наступает, как правило, от остановки дыхания. Например, тетродотоксин, содержащийся в знаменитом японском деликатесе рыбе фугу и некоторых других животных, блокирует пору ПЧНК подобно пробке. Интересно, что тетродотоксин используется чаще в целях защиты. И здесь хорошо исследован пример гонки вооружений: некоторые тритоны используют тетродотоксин для защиты от хищников, а охотящиеся на этих тритонов змеи приобретают устойчивость в результате мутации генов ПЧНК [17]. К блокаторам относятся небольшие пептиды μ-конотоксины, являющиеся оружием морских моллюсков-конусов. Местные анестетики, такие как новокаин и лидокаин, тоже являются блокаторами ПЧНК: их местное применение приводит к блокаде чувствительных нейронов.

Рисунок 3. Некоторые природные нейротоксины, воздействующие на ПЧНК. Изображена модель пространственной организации канала; показан центральный ПД и два ПЧД (II и IV). Представлены фотографии ядовитых животных, служащих источником «классических» нейротоксинов. Тетродотоксин из рыбы фугу блокирует пору ПЧНК, связываясь с внеклеточной стороны. Батрахотоксин из кожи колумбийской лягушки проникает через мембрану и встраивается внутрь поры канала, вызывая его активацию. α-Токсины скорпионов связываются с ПЧД-IV и подавляют процесс естественной инактивации каналов. β-Токсины скорпионов связываются с ПЧД-II и активируют ПЧНК.
Но не все токсины блокируют канал: есть и такие, которые его активируют, то есть увеличивают время, когда канал находится в открытом состоянии. Примером таких молекул является батрахотоксин, содержащийся в секрете кожных желез некоторых Южноамериканских лягушек-листолазов. Распространенные инсектициды пиретроиды также относятся к активаторам ПЧНК.
α- и β-токсины из яда скорпионов представляют собой небольшие белки (~60—70 аминокислотных остатков), в структуре которых присутствует β-лист из трех тяжей и короткая α-спираль, скрепленные четырьмя дисульфидными связями (рис. 4). Хотя и те, и другие токсины, по сути, активируют канал, делают они это по-разному. α-Токсины мешают каналу закрыться, а β-токсины — помогают ему открыться. Соответственно и места взаимодействия с ПЧНК у этих токсинов разные: те и другие связываются с ПЧД, но для α-токсинов это ПЧД-IV, а для β-токсинов — ПЧД-II.

Рисунок 4. Модульная структура α-токсинов скорпионов. Эти токсины — небольшие белки, составленные из β-листа и α-спирали, скрепленных дисульфидными связями. Такая жесткая и эволюционно консервативная структура образует «сердцевину» молекулы, а две петли и C-конец образуют динамически подвижный «модуль специфичности» (показан пунктирным овалом), отличающийся у млеко- (слева) и инсектотоксинов (справа). Рисунок показывает результаты анализа характерных внутренних движений по данным молекулярной динамики. Цветной спектр конформаций изображает динамическую подвижность каждой из молекул.
[23], рисунок модифицирован
Остановимся теперь подробнее на α-токсинах и их «взаимоотношениях» с ПЧНК. Дело в том, что в яде скорпионов присутствуют молекулы, обладающие токсическим действием по отношению к насекомым или млекопитающим (назовем соответствующие α-токсины инсектотоксинами и млекотоксинами). И те, и другие связываются с ПЧД-IV, но только у разных каналов. У насекомых это свой канал, называемый Para [18], а у млекопитающих есть целых девять разновидностей каналов, обозначаемых Nav1.1—1.9 [19]. Разные ПЧНК млекопитающих выполняют разные функции. Например, Nav1.2 характерен для центральной нервной системы, Nav1.4 — для скелетной мускулатуры, Nav1.5 — для сердца [20]. А дальше в ход идет упомянутая «комбинаторная библиотека» яда скорпионов, в которой может найтись молекула, активная по отношению к выбранной мишени. Если это происходит, «владелец» такого яда получает эволюционное преимущество по сравнению с другими, и та часть его генома, которая отвечает за состав ядовитой библиотеки, широко распространяется в будущих поколениях скорпионов.
Присутствующие в яде скорпионов нейротоксины отличаются разнообразием: некоторые из них могут быть направлены исключительно на каналы насекомых (инсектотоксины), тогда как другие действуют на каналы млекопитающих (млекотоксины). Есть также молекулы, действующие сразу и на те, и на другие ПЧНК. Что лежит в основе молекулярной эволюции этих токсинов, позволяющей им угнаться сразу за множеством ускользающих мишеней? Попробуем ответить на этот вопрос.
Компьютерный анализ выявляет «молекулярную гонку вооружений»
В Институте биоорганической химии им. академиков М.М. Шемякина и Ю.А. Овчинникова РАН было проведено исследование, направленное на выявление особенностей млеко- и инсектотоксинов из яда скорпионов, определяющих их селективное взаимодействие с соответствующими ПЧНК. Работа состояла из двух частей:
- Компьютерный анализ структуры и динамики позволил выявить характерные особенности млеко- и инсектотоксинов и предсказать активность «сиротского» токсина с неизвестной селективностью. Эта часть работы была сделана в лаборатории моделирования биомолекулярных систем.
- Затем предсказания были проверены в лаборатории нейрорецепторов и нейрорегуляторов: был синтезирован «сиротский» токсин и исследована его специфичность.
Компьютерное моделирование было основано на методе молекулярной динамики (МД) [21]. Установлено, что молекулы α-токсинов, несмотря на свой небольшой размер и жесткую структуру, состоят из двух динамических модулей. Анализ характерных движений показал, что один из этих модулей достаточно «жесткий», а другой конформационно «пластичный». Более того, движения «пластичного» модуля различаются у млеко- и инсектотоксинов (рис. 4).
Идентифицированные с помощью МД части молекул получили название «сердцевины» и «модуля специфичности». Сердцевина α-токсинов оказывается эволюционно консервативным (очень сходно устроенным у инсекто- и млекотоксинов), а модуль специфичности — вариабельным, соответствующим конкретной мишени действия. Сердцевина, таким образом, отвечает за распознавание ПЧНК «вообще», а быстро изменяющийся в эволюции модуль специфичности позволяет токсину «настраиваться» на конкретный тип канала.
Было также обнаружено, что модуль специфичности млекотоксинов существенно более гидрофильный, чем у инсектотоксинов. Эта особенность, предположительно, отражает структурные детерминанты, позволяющие токсинам селективно распознавать свои мишени. Что интересно, анализ свойств ПЧНК показал ту же тенденцию для внеклеточной части S5—S6 повтора I: у каналов млекопитающих эти области более гидрофильны, а у каналов насекомых — гидрофобны. Сопоставление результатов анализа с накопленными биохимическими данными позволило предложить любопытный характер связывания α-токсинов с ПЧНК. Сердцевинный модуль, по-видимому, взаимодействует с ПЧД-IV, в то время как модуль специфичности связывается с петлей S5—S6 повтора I. Интересно, что данные области в структуре ПЧНК сближены, что также было показано в независимых экспериментах. Идея соответствия модульной организации токсинов доменной структуре каналов имеет интересный эволюционный смысл, позволяющий нам разглядеть гонку вооружений в противостоянии ядовитых животных и их жертв. Модульная структура предположительно позволяет токсинам гибко адаптироваться к изменяющейся мишени.
Компьютерный анализ может быть использован для предсказания активности токсинов с неизученными свойствами. В частности, для токсина M9 из яда Среднеазиатского скорпиона Mesobuthus eupeus, ставшего первым α-нейротоксином из скорпионьего яда, для которого была установлена пространственная структура (кстати, это тоже было сделано в ИБХ [22]), была предсказана активность в отношении каналов как млекопитающих, так и насекомых. Биоинженерный синтез и тестирование активности этого токсина на рекомбинантных ПЧНК подтвердили высказанное предположение. Результаты представленной работы были опубликованы в журнале Journal of Biological Chemistry [23]. Эволюционное обособление модулей α-токсинов, похоже, вызвано требованиями гонки вооружений — необходимостью оперативно адаптироваться вслед за изменяющимися условиями среды: появлением новых мишеней и изменением старых. Вряд ли этот подход является универсальным, но в данном случае он позволил с новой стороны взглянуть на взаимосвязь структура—функция для биологически активных пептидов.
Биоинженерия и нейробиология
Исследования молекулярных основ эволюционной гонки вооружений имеет не только фундаментальное значение. Например, заново созданные «дизайнерские» молекулы — аналоги нейротоксинов с заданной исследователями активностью — являются идеальными инструментами исследования нервной системы. Такие молекулы позволили бы прицельно регулировать работу ионных каналов и модифицировать нервный ответ желаемым образом.
Придание нейротоксинам желаемой селективности и видоспецифичности позволит, например, создать инсектицид нового поколения. Биоинженерно изготовленный или даже внедренный в геном растений инсектотоксин позволит эффективно бороться с насекомыми-вредителями. А можно представить себе возможность создания идеально селективного инсектотоксина, действующего на вредителей и не токсичного для полезных насекомых (например, пчел).
Наконец, идеально селективные нейротоксины — уже не яды, а точно настраиваемые нейрорегуляторы — нашли бы применение в молекулярной медицине для лечения заболеваний, связанных с дисфункцией ионных каналов и называемых каналопатиями. Среди них — различные неврологические и психические расстройства, а также к патологии скелетной и сердечной мускулатуры [24].
Статья поддержана грантом РФФИ д_с-13-04-11520 и первоначально опубликована в «Науке и жизни» [25]. См. также видеозапись «ядовитого» семинара, организованного Советом молодых ученых ИБХ РАН осенью 2013 года.
Видео 1. Доклад А.А. Василевского «Что-то... про паучков», сделанный на «ядовитом» семинаре Совета молодых ученых ИБХ РАН осенью 2013 года. В этом докладе также рассказывается о непростых взаимоотношениях нейротоксинов с их молекулярными мишенями.
Литература
- Докинз Р. Самое грандиозное шоу на Земле. Доказательства эволюции. М.: «Корпус/Астрель», 2012;
- Когда б вы знали, из какого сора…;
- Возможна ли жизнь без гемоглобина?;
- S. G. E. Andersson. (2006). GENETICS: The Bacterial World Gets Smaller. Science. 314, 259-260;
- Семья — женское изобретение;
- Марков А. Эволюция человека. В 2-х томах. М.: «Династия», 2011;
- Элементы: «Гонка вооружений — двигатель эволюции»;
- Элементы: «Современные паразиты опаснее прошлых и будущих»;
- Walter Gassmann, Saikat Bhattacharjee. (2012). Effector-Triggered Immunity Signaling: From Gene-for-Gene Pathways to Protein-Protein Interaction Networks. MPMI. 25, 862-868;
- Иммунологическая Нобелевская премия (2011);
- Толл-подобные рецепторы: от революционной идеи Чарльза Джейнуэя до Нобелевской премии 2011 года;
- Гонки с вирусом: эпидемиология и экология вируса гриппа;
- M. W. van der Woude, A. J. Baumler. (2004). Phase and Antigenic Variation in Bacteria. Clinical Microbiology Reviews. 17, 581-611;
- Великому комбинатору и не снилось: комбинаторика токсинов пауков;
- Формирование мембранного потенциала покоя;
- Frank H Yu, William A Catterall. (2003). . Genome Biol. 4, 207;
- T SOONG, B VENKATESH. (2006). Adaptive evolution of tetrodotoxin resistance in animals. Trends in Genetics. 22, 621-626;
- Ke Dong. (2007). Insect sodium channels and insecticide resistance. Invert Neurosci. 7, 17-30;
- G. F. Lopreato, Y. Lu, A. Southwell, N. S. Atkinson, D. M. Hillis, et. al.. (2001). Evolution and divergence of sodium channel genes in vertebrates. Proceedings of the National Academy of Sciences. 98, 7588-7592;
- W. A. Catterall. (2005). International Union of Pharmacology. XLVII. Nomenclature and Structure-Function Relationships of Voltage-Gated Sodium Channels. Pharmacological Reviews. 57, 397-409;
- Молекулярная динамика биомолекул. Часть I. История полувековой давности;
- Vladimir S. Pashkov, Vladimir N. Maiorov, Vladimir F. Bystrov, Anh N. Hoang, Tatyana M. Volkova, Eugene V. Grishin. (1988). Solution spatial structure of ‘long’ neurotoxin M9 from the scorpion Buthus eupeus by 1H-NMR spectroscopy. Biophysical Chemistry. 31, 121-131;
- Anton O. Chugunov, Anna D. Koromyslova, Antonina A. Berkut, Steve Peigneur, Jan Tytgat, et. al.. (2013). Modular Organization of α-Toxins from Scorpion Venom Mirrors Domain Structure of Their Targets, Sodium Channels. J. Biol. Chem.. 288, 19014-19027;
- G. S.B. Andavan, R. Lemmens-Gruber. (2011). Voltage-Gated Sodium Channels: Mutations, Channelopathies and Targets. CMC. 18, 377-397;
- Чугунов А.О. и Василевский А.А. (2013). Эволюционная «гонка вооружений»: нейротоксины против ионных каналов. «Наука и жизнь». 11, 42–48;
- William A. Catterall. (2010). Ion Channel Voltage Sensors: Structure, Function, and Pathophysiology. Neuron. 67, 915-928;
- О чем не знал Гальвани: пространственная структура натриевого канала;
- Калиевый канал in silico.

Комментарии


