Молекулярная модель синаптической везикулы
18 марта 2007
Молекулярная модель синаптической везикулы
- 1067
- 2
- 9
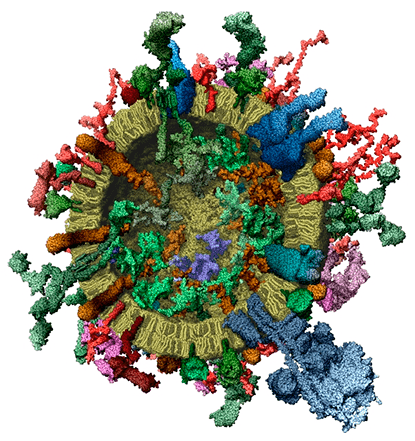
Молекулярная модель усредненной синаптической везикулы
-
Автор
-
Редакторы
Синаптические везикулы осуществляют доставку и высвобождение нейромедиаторов в межсинаптическое пространство в месте соприкосновения двух нервных окончаний, участвуя в передаче сигналов в нервной системе. Недавно опубликованная статья [1] в журнале Cell содержит наиболее подробное на настоящий момент описание синаптической везикулы.
Передача нервного импульса в синапсах осуществляется за счет экзоцитоза (выброса содержимого) синаптических везикул, наполненных нейромедиаторами [2]. Синаптические везикулы (пузырьки) находятся в пресинаптических участках нервных окончаний, причем скапливаются они там в таких больших количествах, что мозг можно рассматривать как микромолярный раствор синаптических везикул. Обилие и одинаковый размер синаптических везикул значительно облегчает их выделение и изучение. Поэтому синаптическая везикула является одной из наиболее полно охарактеризованных органелл в биологии. С момента его первоначального описания, синаптический пузырёк рассматривался как модельная органелла, которая может быть использована для установления общих принципов мембранного транспорта [3]. Эта надежда рано подтвердилась, сразу после открытия белков SNARE, участвующих в слиянии мембран, и белков синаптотагминов, контролирующих запуск слияния при помощи ионов Ca2+. Статья, о которой идет речь — это первое полное количественное описание белкового и липидного состава синаптической везикулы, подтверждающее её статус модельной органеллы.
Ранние работы уже исследовали белково-липидный состав этой органеллы, но не позволяли делать никаких выводов о количественных соотношениях компонентов [4]. Чтобы разрешить этот вопрос, Такамори с коллегами в своей работе использовали комбинацию подходов протеомики (высокопроизводительной белковой химии) и биофизики, что в конце концов позволило им построить первую модель органеллы с атомным разрешением.
Оказалось, что совсем немного видов белков представлены большим количеством копий (синаптобревин: 70 копий на пузырек; синаптофизин: 30 копий; синаптотагмин: 15 копий). Другие же, наоборот, представлены совсем небольшим количеством копий. Наиболее выдающийся пример — протонный насос, одна копия на везикулу. Однако, из-за своего огромного размера, он составляет 10% массы общего белка синаптической везикулы.
Расчет площади поверхности пузырька, занимаемой липидами и трансмембранными участками белковых молекул показал, что последние занимают 20% всей поверхности. Это отношение поразительно велико. Стоит учесть, что трансмембранные участки окружены кольцом неподвижных молекул липидов. Тогда из найденного процентного отношения следует, что в синаптических везикулах очень мало жидких фосфолипидных мембран. Таким образом, белковая составляющая доминирует в этой удивительной органелле, делая её более жёсткой, чем предполагалось.
Литература
- Thomas C. Südhof. (2006). Synaptic Vesicles: An Organelle Comes of Age. Cell. 127, 671-673;
- Katz B.S. The release of neural transmitter substances. Liverpool, UK: Liverpool University Press, 1969. — 70 p.;
- Thomas C. Südhof, Reinhard Jahn. (1991). Proteins of synaptic vesicles involved in exocytosis and membrane recycling. Neuron. 6, 665-677;
- Thomas C. Südhof. (2004). THE SYNAPTIC VESICLE CYCLE. Annu. Rev. Neurosci.. 27, 509-547.

Комментарии


