Из чего же сделан хроматин архей?
09 июля 2019
Из чего же сделан хроматин архей?
- 1779
- 0
- 5
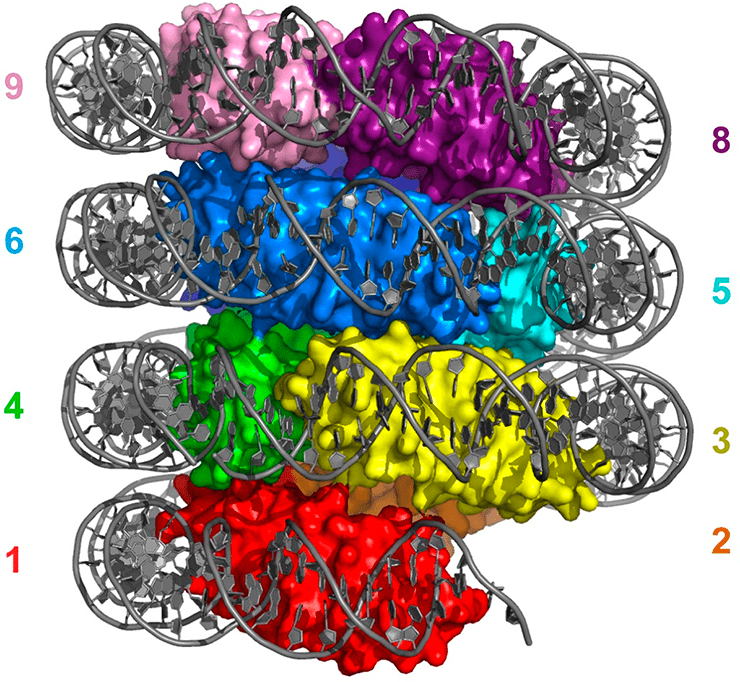
Модель гипернуклеосомы архей на примере гистона HMfB. Димеры гистона формируют левозакрученную спираль, на которую намотана ДНК. Разными цветами отмечены разные димеры гистонов, ДНК закрашена серым.
-
Автор
-
Редактор
Археи, несмотря на то, что не имеют оформленного ядра, по очень многим признакам гораздо больше похожи на эукариот, чем на бактерий. В частности, их геномная ДНК упакована и компактизирована с помощью гистонов, как у эукариот. Однако гистоны эти весьма своеобразны (как, наверное, и всё у архей): в отличие от гистонов эукариот, они не формируют стабильные октамерные нуклеосомы, хотя третичные структуры гистонов архей и эукариот очень похожи. Последние исследования свидетельствуют, что «нуклеосомы» архей не имеют фиксированного размера и состоят из различного числа димеров гистонов, причем плотность упаковки ДНК с помощью таких вариабельных нуклеосом напрямую связана с репрессией транскрипции связанного с ними участка ДНК. Что наиболее удивительно, длина нуклеосом архей, похоже, может быть практически неограниченной, за что исследователи назвали их гипернуклеосомами. Впрочем, с помощью биоинформатического анализа у некоторых архей удалось найти гистоны с сильно отличающейся от остальных аминокислотной последовательностью, которые, по-видимому, неспособны формировать гипернуклеосомы. Наконец, у некоторых архей есть гистоны с N- и C-концевыми хвостами, которые похожи на хвосты гистонов эукариот и тоже могут подвергаться посттрансляционным модификациям. Так каковы же они, гистоны архей, и как устроен хроматин архей? В статье мы постараемся ответить на эти вопросы.
Необходимость поддерживать пространственную архитектуру хроматина стоит перед представителями всех трех доменов жизни: архей, бактерий и эукариот. У бактерий имеются особые белки — нуклеоид-ассоциированные белки (nucleoid-associated proteins, NAPs), позволяющие изгибать или переплетать спирали ДНК, и эти белки не только определяют структуру бактериального хроматина, но и влияют на активность определенных участков генома. К числу таких белков относятся, в частности, гистоноподобный белок HU из штамма U93 E. coli, а также H-NS — гистоноподобный белок, структурирующий нуклеоид. У эукариот и большинства архей задачу упаковки геномной ДНК выполняют гистоны [1]. Хотя наша статья посвящена гистонам архей, будет нелишним напомнить, как устроены гистоны эукариот.
Ноу-хау архей: бесконечная гипернуклеосома
Что же касается архей, то гистоны имеются у большинства видов в самых разных группах. Любопытно, что у некоторых есть и гомолог бактериального гистоноподобного белка HU, а у многих архей идентифицированы особые высококонсервативные нуклеоид-ассоциированные белки семейства Alba, которые, впрочем, найдены и у эукариот (рис. 2). Эти белки способны к формированию филаментов при взаимодействии с ДНК, а также соединяют дуплексы ДНК подобно мостикам. Эксперименты в условиях in vitro показали, что связывание ДНК с белками Alba подавляет транскрипцию. Любопытно, что у большинства архей типа кренархеотов отсутствуют гистоны, и ведущую роль в организации хроматина играют NAPs [1].

Рисунок 2. Модель организации хроматина архей
Существуют и другие, более малочисленные белки, влияющие на упаковку ДНК, которые содержат некоторые кренархеоты и эвриархеоты. Например, у кренархеотов имеются специфические мономерные белки, изгибающие ДНК, — Cren7 и Sul7 [1].
Гистоны архей, как и гистоны эукариот, имеют гистоновую складку, однако, как правило, лишены N-концевых хвостов. Не выявлено у архей и линкерных гистонов, гомологичных эукариотическому гистону H1.
В растворе гистоны архей образуют димеры — как гомодимеры, так и гетеродимеры, поскольку многие археи синтезируют гистоны нескольких видов. К примеру, архея Methanothermus fervidus производит два разных гистона, уровень синтеза и относительное соотношение которых различны в разных фазах роста, поэтому, возможно, эти белки имеют разное функциональное значение [1].
Имеются сведения, что как in vitro, так и in vivo гистоны архей могут связывать ДНК в виде тетрамеров. Ряд исследований показал, что в действительности ДНК архей связана с неограниченным числом димеров гистонов, которые, взаимодействуя друг с другом, формируют ДНК-белковый филамент вариабельной, теоретически неограниченной длины. Кристаллографическое изучение гистона HMfB вышеупомянутой археи M. fervidus показало, что in vitro его молекулы формируют бесконечную левозакрученную нить — гипернуклеосому, — причем чем больше витков в ней, тем плотнее упакована намотанная на нее ДНК. Когда структуру гипернуклеосомы у археи Thermococcus kodakarensis экспериментально нарушали, вводя мутации в гистоны, архея начинала иначе реагировать на определенные питательные вещества, что свидетельствует о роли упаковки ДНК архей в экспрессии генов [1].
Изучая гистон HMfB, исследователи выяснили, что он образует димеры за счет гидрофобных взаимодействий аминокислотных остатков, расположенных в глубине молекулы, а также посредством гистоновых складок, для формирования которых необходим один крайне важный солевой мостик. Эти гидрофобные остатки, а также остатки, образующие солевой мостик, оказались высококонсервативными среди гистонов архей, поэтому гистоны архей, вероятнее всего, в большинстве своем имеют схожую структуру. Кроме того, консервативными оказались и аминокислоты, которые обеспечивают взаимодействие гистонов архей с малой бороздкой ДНК. На основании перечисленных данных высказали предположение, что гипернуклеосомы характерны для хроматина большинства архей. Впрочем, некоторые новооткрытые археи из прежде неизвестных типов имеют крайне необычные гистоны, о которых расскажем ниже [1].
Эволюция гистонов
Предполагается, что эволюционно гистоны эукариот произошли от гистонов архей. Родство этих белков подтверждается высоким сходством и аминокислотных последовательностей, и вторичных структур. По расчетам исследователей, первые гистоны появились более одного миллиарда лет назад именно как структурные белки, а участвовать в регуляции экспрессии генов они стали гораздо позже, когда у эукариот обособились четыре типа коровых гистонов [2]. Интересно, что гистоновая складка есть не только у некоторых эукариотических белков, не являющихся гистонами (таких как некоторые факторы транскрипции), но и у белков некоторых бактерий. Биоинформатический анализ последовательностей этих белков показал, что мотив гистоновой складки существовал уже во времена последнего универсального общего предка (LUCA) [3].
Подробности о том, как выглядел LUCA, и как ему жилось на доисторической Земле, читайте в статьях: «В диких условиях: как жил последний всеобщий предок LUCA» [4] и «От Бульона до Эукариот. Первый организм и наш древнейший предок» [5]. — Ред.
Как упоминалось выше, элементарная единица комплексов гистонов большинства архей — димер, но у архей класса Halobacteria имеются необычные тандемные гистоны. Эти гистоны представляют собой сшитый в одну полипептидную цепь димер с двумя взаимодействующими гистоновыми складками [2], что подчеркивает важность и необходимость гистоновой складки для формирования такого димера. Вместе с тем давление отбора на остальную часть молекулы тандемного гистона ослабевало, и, согласно одной из гипотез, из тандемного гистона в конце концов появился асимметричный гистон-димер, который мог стать предком эукариотического димера H3—H4. Впоследствии асимметричный гистон разъединился на два разных белка, кодируемых двумя разными генами. Действительно, тетрамер гистонов архей структурно очень близок к эукариотическому тетрамеру H3—H4, а гистоны H2A и H2B, как предполагается, произошли позже от разделившихся гистонов H3 и H4. В пользу такого сценария свидетельствует и тот факт, что гистоны H3 и H4 гораздо больше похожи на гистоны архей, чем гистоны H2A и H2B [1].
Далее у эукариот начали появляться так называемые неканонические формы гистонов, которые часто приурочены к определенным клеточным процессам, причем, что интересно, неканонических форм гистонов H2B и H4 практически неизвестно, в то время как для H3 и H2A они описаны в изобилии. Дальнейшим усложнением эукариотических гистонов стало формирование октамеров, на которые ДНК наматывается в виде двух витков, а не одного, а также появление N-концевых хвостов, которые позволили еще плотнее компактизировать ДНК и стали подвергаться интенсивным посттрансляционным модификациям [1].
Но наиболее интересно обстоит дело с гистонами у новооткрытых архей, которые стали известны науке благодаря бурному развитию метагеномики . Например, в собранном геноме LC_3 предполагаемого типа Heimdallarchaeota удалось найти гены целых десяти гистонов — абсолютный рекорд для архей! Вместе с тем у представителей предполагаемых типов Parvarchaeota, Geothermarchaeota и Vestraetearchaeota гистонов не нашлось вовсе: у этих архей, по-видимому, за компактизацию хроматина отвечают NAPs, тем более что у некоторых из них удалось найти упаковывающие белки вышеупомянутого семейства Alba. Похожая ситуация и в случае относительно давно описанных кренархеот, подавляющее большинство которых не имеет гистонов. В геномах Geothermarchaeota и Vestraetearchaeota исследователи пока не смогли найти генов, белковые продукты которых походили бы на известные белки хроматина. Наконец, у представителей предполагаемого типа Thorarchaeota есть ген, кодирующий белок HU (как мы помним, он отвечает за упаковку ДНК в нуклеоидах кишечной палочки) [1].
О метагеномике и эволюции архей рассказывает статья «Закинули археи эволюционный невод и вытянули...» [6]. — Ред.
Посттрансляционные модификации гистонов архей
Долгое время было принято считать, что гистоны архей не подвергаются посттрансляционным модификациям, а их регуляторная роль сводится к блокировке транскрипции ДНК, с которой они связаны, за счет того, что они не дают РНК-полимеразе II связываться с ДНК и синтезировать РНК-транскрипт. Так, у археи T. kodakarensis, имеющей два разных гистона, делеция одного из них приводила к почти десятикратному усилению экспрессии около 5% генов (однако делеция обоих гистонов оказалась несовместима с жизнью, что указывает на важнейшую роль гистонов в функционировании клетки). Ранние эксперименты по транскрипции in vitro с транскрипционной машинерией археи Methanothermobacter thermoautotrophicus показали, что добавление гистонов к раствору с ДНК и белками транскрипции снижало количество вырабатываемых транскриптов, а при добавлении гистонов в концентрации, в которой они, согласно расчетам, должны связывать всю находящуюся в растворе ДНК (около 1 димера гистонов на 30 пар оснований ДНК) синтез РНК останавливался вовсе. Схожие данные были получены для транскрипционного аппарата Pyrococcus furiosus.
Считается, что архейные транскрипционные факторы TBP и TFB способствуют транскрипции, поскольку конкурируют с гистоновыми димерами за связывание с ДНК и в конечном счете вытесняют их (рис. 3а). У T. kodakarensis удалось выявить, что фактор транскрипции TFS, а также комплекс Spt4/5 облегчает взаимодействие РНК-полимеразы с ДНК, что также активирует экспрессию генов (рис. 3б) [2].

Рисунок 3. У архей транскрипционные факторы вытесняют димеры гистонов с ДНК и тем самым способствуют активации экспрессии генов
Новооткрытые группы архей принесли биологам и другие сюрпризы. Оказалось, что некоторые представители предполагаемых типов Heimdallarchaeota, Huberarchaea и Bathyarchaeota имеют гистоны с N-концевыми хвостами, как гистоны эукариот! Хвосты двух гистонов, найденных у Heimdallarchaeota и Huberarchaea, имеют примерно ту же длину и аминокислотный состав, что и хвост гистона H4. Поскольку четыре остатка лизина в N-концевом хвосте H4 подвергаются ацетилированию, гипотетически они могут ацетилироваться и в хвостах гистонов архей, тем более что в их геномах найдено несколько предполагаемых лизинацетилтрансфераз и деацетилаз. Более того, идентифицировали даже белок, который может распознавать эти ацетилированные остатки. Так что археи, вполне вероятно, используют посттрансляционные модификации гистонов для регуляции уровня компактизации генома и экспрессии генов, как эукариоты. Более того, некоторые гистоны архей имеют не N-концевые, а C-концевые хвосты, похожие на короткий C-концевой хвост эукариотического гистона H4. В геномах некоторых архей найдены гены, кодирующие «усеченные» версии гистонов, которые, тем не менее, способны связываться с ДНК и выполнять некоторые регуляторные функции [1]. Показано также, что посттрансляционным модификациям могут подвергаться и NAPs семейства Alba [2].
Несмотря на значительный прогресс в нашем понимании устройства хроматина архей, многое еще предстоит выяснить. Неизвестно, например, насколько динамичны гипернуклеосомы, и как клетки регулируют размер теоретически бесконечных гипернуклеосом? Как на структуре гипернуклеосомы отражается синтез особых форм гистонов в определенных условиях [1]? Остается надеяться, что дальнейшее изучение «темной материи» хроматина архей поможет ответить на эти и многие другие вопросы.
Литература
- Bram Henneman, Clara van Emmerik, Hugo van Ingen, Remus T. Dame. (2018). Structure and function of archaeal histones. PLoS Genet. 14, e1007582;
- Travis J. Sanders, Craig J. Marshall, Thomas J. Santangelo. (2019). The Role of Archaeal Chromatin in Transcription. Journal of Molecular Biology;
- Vikram Alva, Andrei N Lupas. (2019). Histones predate the split between bacteria and archaea. Bioinformatics. 35, 2349-2353;
- В диких условиях: как жил последний всеобщий предок LUCA;
- От Бульона до Эукариот. Первый организм и наш древнейший предок;
- Закинули археи эволюционный невод и вытянули....

Комментарии


